Architektur von Blütenpflanzen
Forschungsbericht (importiert) 2006 - MPI für Pflanzenzüchtungsforschung
Morphogenese höherer Pflanzen
Grundlagen der Formenvielfalt von Pflanzen sind in hohem Umfang die Verzweigungsmuster des Sprosses und der Blütenstände, die durch die Anzahl, Anordnung und Entwicklungsintensität der Seitentriebe geprägt werden. Seitentriebe werden gebildet aus sekundären Meristemen (Achselmeristeme oder Lateralmeristeme), das sind Gruppen teilungsfähiger Zellen, die sich in den Achseln von Blättern entwickeln. Die Achselmeristeme initiieren die Bildung von Blattanlagen (Blattprimordien), was schließlich zur Entstehung einer Knospe führt. Solche Achselknospen können entweder für unbestimmte Zeit ruhen oder aber zu einem Seitentrieb auswachsen. Die Unterdrückung des Auswachsens geht auf einen inhibitorischen Effekt der primären Sprossspitze zurück, der als Apikaldominanz bezeichnet wird. Dabei üben die Pflanzenhormone Auxin und Cytokinin gegensätzliche Wirkungen aus: Auxin unterdrückt das Auswachsen der Achselknospen, während Cytokinin dies fördert. Die Ausbildung ruhender Achselknospen ist ein wichtiges Element im Entwicklungszyklus mehrjähriger Wuchsformen (zum Beispiel bei Bäumen).
In den Blütenständen entwickeln sich die Lateralmeristeme zu Blüten und haben somit eine entscheidende Funktion für den Erhalt und die Ausbreitung einer Art. Im Laufe der letzten Jahrtausende hat die Nutzbarmachung von Pflanzen durch den Menschen zu einer Anpassung der Wuchsformen an die Anforderungen der Landwirtschaft und des Gartenbaus geführt. In der Landwirtschaft werden häufig unverzweigte Wuchsformen bevorzugt, die zu einer Konzentration der pflanzlichen Ressourcen in wenigen, ertragreichen Fruchtständen und einer leichten Handhabbarkeit führen. Ein bekanntes Beispiel dafür ist die Domestizierung des Mais aus der Wildform Teosinte, bei der eine Veränderung in einem einzigen Gen eine deutliche Erhöhung des Ertrags zur Folge hatte [1]. Im Gegensatz dazu wird bei Zierpflanzen häufig ein stark verzweigtes Sprosssystem bevorzugt, da dies eine Vielzahl von Blüten garantiert.
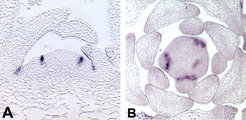
Forscher der Abteilung Pflanzenzüchtung und Genetik am Max-Planck-Institut für Züchtungsforschung sind daran interessiert, die genetischen Mechanismen zu verstehen, die die Verzweigung des Sprosssystems kontrollieren. Dabei konzentrieren sich die Arbeiten auf den ersten entscheidenden Schritt, nämlich die Anlage von neuen Meristemen in den Achseln der Blätter. Bis heute ist nicht bekannt, wie diese Achselmeristeme entstehen. Eine Arbeitshypothese besagt, dass schon bei der Anlage eines Blattprimordiums eine Gruppe teilungsfähiger Zellen aus dem Hauptmeristem der Sprossspitze abgegliedert und in den Achseln der Blattanlagen deponiert wird [2]. Diese Gründerzellen könnten sich später zu einem neuen Meristem entwickeln. Alternativ könnten Achselmeristeme aus bereits teilweise oder vollständig differenzierten Zellen entstehen. In verschiedenen Pflanzenarten (zum Beispiel Tomate, Löwenmäulchen oder Erbse) sind Mutanten beschrieben worden, die die Fähigkeit zur Bildung von Seitentrieben verloren haben. Bei der Tomatenmutante lateral suppressor (ls) ist die Bildung von Achseltrieben in der vegetativen Entwicklungsphase stark reduziert (Abb. 1). Allerdings zeigt diese Mutante keine Veränderung im Verzweigungsmuster des Blütenstandes.
 und der lateral suppressor-Mutante (B) in Tomate.")

Genetische Kontrolle des vegetativen Wachstums
Vor einigen Jahren konnte gezeigt werden, dass der Verzweigungsdefekt in der ls-Mutante darauf zurückzuführen ist, dass ein bestimmtes Protein fehlt: Das Ls-Protein fungiert, so die Annahme, sehr wahrscheinlich als Transkriptionsfaktor [3]. Transkriptionsfaktoren sind Proteine, die an bestimmten Stellen der DNA, den so genannten Promotoren, binden und die Expression von dahinter (stromabwärts der genetischen Information) oder in der Nähe gelegenen Genen kontrollieren. Inzwischen konnte gezeigt werden, dass Arabidopsis [4] und Reis [5] ähnliche Gene besitzen. Das LATERAL SUPPRESSOR-Gen aus Arabidopsis (LAS) kann nach einer Übertragung in die ls-Mutante der Tomate die dort fehlende Funktion ersetzen [4], woraus gefolgert werden kann, dass das LAS-Protein eine Schlüsselfunktion besitzt, die über große evolutionäre Distanzen konserviert ist. Umso interessanter ist die Frage, welche Rolle das LAS-Protein bei der Anlage von Achselmeristemen spielt. Dazu wurde zunächst untersucht, in welchen Zellen die mRNA des LAS-Gens zu finden ist. In Gewebeschnitten der Sprossspitze von Arabidopsis konnte die LAS-mRNA in wenigen Zellen der Blattachsel nachgewiesen werden (Abb. 2). In den umgebenden Geweben ist die LAS-mRNA dagegen nicht zu finden.
 und ein Querschnitt (B) durch die Sprossspitze wurden mit einer Sonde für das LATERAL SUPPRESSOR-Gen hybridisiert. Die dunkelblau gefärbten Zellen enthalten LAS-mRNA.")
Achsel-Zellen, die LAS exprimieren, unterscheiden sich von den umgebenden Zellen auch dadurch, dass sie cytoplasmareich sind und kaum Vakuolen besitzen. LAS-mRNA wurde in den Achseln der Blattprimordien gefunden, ehe die Anlage eines Achselmeristems nachweisbar war, was darauf hinweist, dass die LAS-Aktivität vor der Anlage des Achselmeristems benötigt wird. Vermutlich ist das LAS-Protein daran beteiligt, die Kompetenz der Gründerzellen für eine Meristembildung aufrecht zu erhalten. Dieses Ergebnis unterstützt die eingangs beschriebene Hypothese, nach der Achselmeristeme aus Gründerzellen entstehen, die vom Hauptmeristem der Sprossspitze rekrutiert wurden und ihre Kompetenz zur Meristembildung behielten.
Feinregulation der Sprossarchitektur
Die Charakterisierung eines weiteren Gens, das die Sprossverzweigung reguliert, geht zurück auf eine Tomatenmutante, genannt blind, die Störungen in der Verzweigung des Sprosses und der Infloreszenz aufweist [6]. Das Blind-Gen kodiert einen MYB-Transkriptionsfaktor, der, wie im Falle von Ls, die Expression von anderen Genen kontrolliert. Sowohl in Tomate als auch in Arabidopsis gibt es eine kleine Gruppe von verwandten MYB-Genen, die vermutlich ähnliche Funktionen besitzen. Die Charakterisierung von Mutationen innerhalb dieser Genfamilie in Arabidopsis zeigte, dass einzelne Gene (RAX1, RAX2, RAX3), allein oder in Kombination, die Anlage von Achselmeristemen in unterschiedlichen Zonen entlang der Sprossachse kontrollieren [7]. Auch die RAX-Gene werden, wie Ls, zum Teil (RAX1 und RAX3) in eng begrenzten Bereichen der Blattachsel exprimiert. Interessant ist in diesem Zusammenhang auch, dass der Verlust der Genfunktionen von RAX1-RAX3 nur dann zu Störungen in der Verzweigung führt, wenn die Pflanzen im Kurztag, also mit acht Stunden Licht, angezogen wurden. Unter Langtag-Bedingungen (16 Stunden Licht) scheint die Pflanze andere Gene zu aktivieren, die den Verlust der RAX-Genfunktionen ersetzen können. Diesen aus evolutionärer Sicht interessanten Kontrollmechanismus nutzen vielleicht verschiedene Pflanzenarten für eine Feinabstimmung der Achselmeristemanlage und somit auch der Sprossverzweigung.
Signale aus der Wurzel
Das Auswachsen der Achselknospen wird durch Umweltbedingungen und hormonelle Signale gesteuert. Bisher ging man davon aus, dass der Knospenaustrieb im Wesentlichen durch die Pflanzenhormone Auxin und Cytokinin reguliert wird. Das in der Sprossspitze gebildete Auxin hat einen hemmenden Einfluss auf den Austrieb der Knospen, während in der Wurzel synthetisiertes Cytokinin das Auswachsen fördert [8]. Neuere Arbeiten haben ein weiteres Signal nachgewiesen, das den Austrieb von Achselknospen unterdrückt. Dieses Signal wird, wie Pfropfungsexperimente gezeigt haben, in der Wurzel generiert, gelangt von dort in den Spross und unterdrückt das Auswachsen der Achselknospen. In Arabidopsis wurden vier Gene identifiziert (MAX1-MAX4), die an der Synthese, dem Transport und der Perzeption dieses beweglichen inhibitorischen Signalmoleküls beteiligt sind [9]. Zurzeit wird versucht, die chemische Struktur dieses Inhibitors aufzuklären.