Blütenbildung und Fruchtbarkeit: Wird sie von weiteren noch unbekannten Genen außer von den MADS-box Genen gesteuert?
Forschungsbericht (importiert) 2003 - MPI für Pflanzenzüchtungsforschung
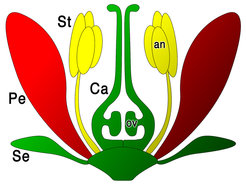
Das beste Unterscheidungsmerkmal von Höheren Pflanzen sind die Blüten. Man nennt sie deshalb Blütenpflanzen. Blüten sind sehr komplexe Organe, die aus unterschiedlichen Organen (Abb. 1), wie zum Beispiel Kelch- (Se), Blüten- (Pe), Staub (St) und Fruchtblättern (Ca) bestehen. Vor mehr als zehn Jahren wurden Gene entdeckt, die eine große Rolle bei der Bildung der einzelnen Blütenteile spielen. Es sind die MADS-box-Gene. Sie enthalten die Bauanleitung für bestimmte Eiweiße (Proteine), in diesem Falle Transkriptionsfaktoren, die für die Initiation wichtiger molekularbiologischer Prozesse essentiell sind [1, 2]. Mutationen der MADS-box-Gene, also Veränderungen der Erbinformation, führte zu Fehlbildungen der Blüten. Man beobachtete z. B. Blüten, die anstelle der Blütenblätter weitere Kelchblätter ausbildeten. Seit der Entdeckung der MADS-box-Gene forschen die Wissenschaftler in meiner Arbeitsgruppe (Abteilung Molekulare Pflanzengenetik am Max-Planck-Institut für Züchtungsforschung in Köln) an folgender Fragestellung: "Welche Rolle spielen die MADS-box-Gene bei der Blütenbildung und in der Evolution der Blütenpflanzen?"
, wie zum Beispiel Kelch- (Se), Blüten- (Pe), Staub (St) und Fruchtblättern (Ca) bestehen.")
Einige Pflanzen haben wunderschöne Blüten, leckere Früchte oder Samen. Am wichtigsten ist es aber für die Pflanze, dass sie sich fortpflanzen kann und somit ihr Überleben sichert.
Geschlechtszellen (Keimzellen) oder Gameten werden in einem Prozess gebildet, bei dem eine meiotische Reduktionsteilung stattfindet, die den diploiden zu einem haploiden Chromosomensatz reduziert. Bei Tieren werden die Keimzellen, die Ei- bzw. Spermazellen, bei der Entwicklung des Lebewesens schon im Embryonalstadium von den sich differenzierenden Körperzellen getrennt. Bei den Pflanzen entstehen die Keimzellen erst viel später in der Entwicklung in den Blüten der Pflanze. Die Samenpflanzen durchlaufen einen Generationswechsel: Es wechseln sich - wie bei Farnen und Moosen - Sporophyt und Gametophyt ab. Der Sporophyt ist die Pflanze, die gemeinhin als Baum, Strauch oder Kraut bekannt ist. Die Fortpflanzungsorgane befinden sich in der Blüte. Dort werden in den Pollensäcken der Staubblätter die Pollenkörner erzeugt. In den Fruchtblättern bildet sich der Embryosack. Der männliche Gametophyt wächst aus dem Pollenkorn (das den Sporen eines Farnes entspricht) als Pollenschlauch heraus, der durch den Griffel zum Embryosack wächst und dort die Eizelle befruchtet. Der Embryosack stellt den weiblichen Gametophyten dar. Er bildet die Eizelle. Beide Gametophyten haben nur den halben Chromosomensatz des Sporophyten.
SBP-box-Gene: eine pflanzeneigene Gruppe von Transkriptionsfaktoren
Auf der Suche nach Steuerelementen für die MADS-box-Gene, entdeckten wir vor einigen Jahren eine neue Familie von Transkriptionsfaktoren. Einige von ihnen stehen für Proteine, die an definierten Stellen der DNA in der Promoterregion der Antirrhinum-MADS-box-Gene namens "SQUAMOSA"[3] binden und deshalb SQUAMOSA-Promotor-Binde-Proteine (SBP) genannt werden [4]. Der Promotor ist die Startstelle auf der DNA, ab welcher das Polymerase-Enzym eine Kopie der DNA in Form einer messenger-RNA (mRNA) herstellen kann, die für die Produktion von Proteinen in der Zelle unerlässlich ist.
Transkriptionsfaktoren werden für die wichtigsten Regulatoren in Entwicklungsprozessen gehalten. Wir wollten wissen, welche Aufgaben sie in Pflanzen erfüllen. Bei einem Experiment mit genetisch veränderten Arabidopsis-Pflanzen stellet sich heraus, dass eines der ersten SPB-box-Gene das frühzeitige Blühen hervorruft [5].
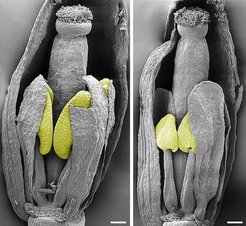
Trotz des Versuches, die Funktion der SBP-Transkriptionsfaktoren während des Pflanzenwachstums mithilfe transgener Arabidopsis-Pflanzen zu entschlüsseln, blieb deren genaue Aufgabe immer noch unklar. Aus diesem Grunde schalteten wir die SBP-Gene aus und erforschten die Auswirkungen des Fehlens der verschiedenen SBP-Transkriptionsfaktoren in der Pflanze. Mit diesem Experiment erhielten wir folgende interessante Ergebnisse [6]: Ein SBP-box-Gen beeinflusst die Sporogenese. Das Gen ist in Wissenschaftlerkreisen unter dem Namen SPL8 bekannt und ist eines von 16 SQUAMOSA-Promotor-Binde-Protein-ähnlichen Genen [7], die man in der Ackerschmalwand (Arabidopsis) gefunden hat. Die spl8-Pflanzenmutanten kann man erst voneinander unterscheiden, wenn sie blühen. Die Mutanten entwickeln keine Samenanlage mit den typischen verlängerten Fruchtblättern und haben deshalb auch keine Samen (Abb. 2).


Auf rasterelektronenmikroskopischen Aufnahmen sieht man noch weitere Missbildungen der SPL8-Mutanten (Abb. 3).
 mit der des Wildtyps kurz vor ihrem Öffnen sieht man den sehr ähnlichern Aufbau. Auf den Fruchtblättern sitzen die pinselartigen Papillae, über die der Pollen nach dem Öffnen der Blüte empfangen wird. Auch sind die Größen der Blüten- und Kelchblätter vergleichbar. Für die Aufnahme wurden einige entfernt. Trotzdem erkennt man auch einen eindeutigen Unterschied in der Größe der Staubbeutel (gelb) zwischen Wildtyp (links) und spl8-Mutante (rechts). Zum Größenvergleich der 100 µm lange weiße Strich.")
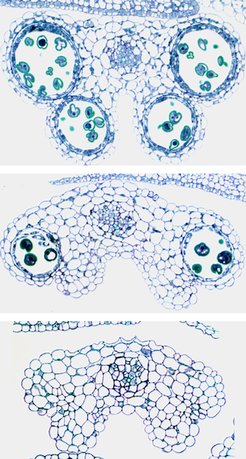
Analysen des Pflanzengewebes zeigten, dass diese Verformungen durch unnormale Entwicklung der vier Mikrosporangien, die später die vier Pollensäcke eines Staubblattes bilden, entstehen. Die Pollensäcke sind viel kleiner als bei normalen Pflanzen (Abb. 4) oder sie fehlen sogar ganz.
 sieht man alle vier Pollensäcke. Bei den Schnitten durch die Pollensäcke der spl8-Mutante kann man sehen, dass sich entweder nur einige kleine Pollensäcke mit weniger Pollen entwickeln (Mitte) oder gar keine entstehen (unten).")
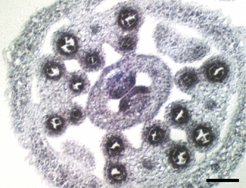
Die Fehlbildung des Gewebes, das man in den Staubblättern der spl8-Mutanten untersucht hat, steht mit der Aktivität der Gene in diesen Gewebeteilen in enger Verbindung (Abb. 5).
 kann man die Expression von SPL8 in den Zellschichten der vier Pollensäcke in allen Staubbeuteln durch das molekularbiologische Verfahren der In-situ-Hybridisierung nachweisen. Zum Größenvergleich der 50 µm lange weiße Strich.")
Unsere Analysen und Untersuchungen der spl8- Mutanten beweisen, dass das SPL8-Gen für die richtige Entwicklung der Geschlechtszellen von Pflanzen eine sehr wichtige Rolle spielt. Bemerkenswerterweise ist das SPL8-Gen nicht für die Bildung der Geschlechtszellen essentiell. Die Mutanten konnten immer noch Pollen und Samen, wenn auch in sehr geringem Umfang, bilden. Aus dieser Beobachtung schließen wir, dass das SPL8-Gen in einem komplexen Kontrollmechanismus bei der Entwicklung und Bildung von in den Staubblättern und somit für die Bildung von Pollen beteiligt ist. Wir vermuten, dass auch das Pflanzenhormon Gibberellinsäure ein Baustein dieses Kontrollmechanismus sein könnte. Mutanten, die sensibel auf Gibberellinsäure reagieren, zeigten häufig Probleme bei der Fortpflanzungsfähigkeit.
Zusammenfassung
Ohne Blüten können auch keine Samen entstehen und die Pflanze kann sich nicht fortpflanzen. Die Blütenpflanzen mussten sich im Laufe der Evolution an ganz unterschiedliche Umweltbedingungen anpassen. Aus diesem Grunde gibt es große Unterschiede im Blühzeitpunkt, im Bau der Blüte und in der Art und Weise der Bestäubung. Die MADS-box-Transkriptionsfaktoren sind ausschlaggebende Regulatoren bei wichtigen Fragen: Wann, wo und wie werden die Blüten gebildet. Trotzdem sind noch weitere Regulationsfaktoren notwendig, um Gewebedifferenzierung in den Blüten zu steuern. Einer davon ist das SBP-box-Gen SPL8. Es gehört zu einer Gruppe von pflanzeneigenen Transkriptionsfaktoren. In einer weiteren Untersuchung und Erforschung der Funktionen des SPL8-Gens wollen wir zur Aufklärung der Gewebedifferenzierung in den Blütenorganen beitragen, die zur Bildung der Geschlechtszellen in Pflanzen führen.
Danksagung:
Ein Teil dieser Forschungsarbeit wurde durch die Deutsche Forschungsgemeinschaft (DFG) unterstützt.
Literatur:
1. Sommer, H., Beltran, J.-P., Huijser, P., Pape, H., Lönnig, W.-E., Saedler, H., Schwarz-Sommer, Zs. (1990). Deficiens, a homeotic gene involved in the control of flower morphogenesis in Antirrhinum majus: the protein shows homology to transcription factors. EMBO J. 9:605-613.
2. Schwarz-Sommer, Zs., Huijser, P., Nacken, W., Saedler, H., Sommer, H. (1990). Genetic control of flower development by homeotic genes in Antirrhinum majus. Science 250:931-936.
3. Huijser, P., Klein, J., Lönnig, W. E., Meijer, H., Saedler, H., and Sommer, H. (1992) Bracteomania, an inflorescence anomaly, is caused by the loss of function of the MADS-box gene SQUAMOSA in Antirrhinum majus. EMBO J. 11, 1239-1249.
4. Klein, J., Saedler, H., and Huijser, P. (1996). A new family of DNA binding proteins includes putative transcriptional regulators of the Antirrhinum majus floral meristem identity gene SQUAMOSA. Mol. Gen. Genet. 250, 7-16.
5. Cardon, G.H., Höhmann, S., Nettesheim, K., Saedler, H., and Huijser, P. (1997). Functional analysis of the Arabidopsis thaliana SBP-box gene SPL3: a novel gene involved in the floral transition. Plant J. 12, 367-377.
6. Unte, U.S., Sorensen, A.-M., Pesaresi, P., Gandikota, M., Leister, D., Saedler, H., Huijser, P. (2003). SPL8, a SBP-Box Gene Affecting Pollen Sac Development in Arabidopsis. Plant Cell 15:1009-1019.
7. Cardon, G.H., Höhmann, S., Klein, J., Nettesheim, K., Saedler, H., and Huijser, P. (1999). Molecular characterisation of the Arabidopsis SBP-box genes. Gene 237, 91-104.