Die Entstehung eines robusten Immunsystems bei Pflanzen
Forschungsbericht (importiert) 2011 - MPI für Pflanzenzüchtungsforschung
Immunabwehr ist Teil des Stressnetzwerks
Pflanzenkrankheiten können große Verwüstungen hinterlassen. Trotzdem sind die meisten Pflanzen sehr gut in der Lage, Infektionsversuche abzuwehren. Nur diejenigen Stämme, die das Abwehrsystem der Pflanzen ausschalten können, sind erfolgreiche Krankheitserreger. Pflanzen haben im Gegenzug dazu ein zelluläres Netzwerk zur Bewältigung von Stress entwickelt, mit dem sie die Eindringlinge bekämpfen. Jetzt geht es darum zu verstehen, welche Dynamik bei der Übermittelung der Resistenzsignale auftritt, wie bestimmte Stoffwechselwege funktionieren, welche Resistenzleistungen tatsächlich notwendig sind und wie die verschiedenen Stoffwechselwege zur Stressbewältigung miteinander kommunizieren.
Verschiedene Ebenen der angeborenen Immunität
Vieles von dem, was über die Immunabwehr der Pflanzen bekannt ist, stammt aus Untersuchungen an der Modellpflanze Arabidopsis thaliana. Dabei wurden fundamentale Prinzipien entdeckt. So ist ein Bild der pflanzlichen Immunabwehr entstanden, mit dem auch die Erforschung von Krankheiten bei komplexeren und genetisch weniger zugänglichen Kultur- und Nutzpflanzen angegangen werden kann. Bisher sieht es so aus, dass die gefundenen regulatorischen Prinzipien überall im Pflanzenreich gleich sind. Pflanzen entwickeln sich parallel zu ihren krankmachenden Feinden in aufeinanderfolgenden Zyklen von Angriff und Gegenangriff [1].
, die von dem Immunregulator EDS1 abhängig ist: Ist dieser in den Pflanzen nicht aktiv (rechtes Bild), bleibt die Autoimmunreaktion aus und die Pflanzen zeigen normales Wachstum.Die falsche Aktivierung eines Rezeptors bewirkt eine Autoimmunreaktion mit der Folge eines stark verlangsamten Wachstums der Pflanzen. Zu sehen ist die durch massive Akkumulation und dadurch Autoaktivierung des RPS4 Rezeptorproteins bedingte Autoimmunität bei Arabidopsis (linkes Bild), die von dem Immunregulator EDS1 abhängig ist: Ist dieser in den Pflanzen nicht aktiv (rechtes Bild), bleibt die Autoimmunreaktion aus und die Pflanzen zeigen normales Wachstum.")

Die stärkste Reaktion eines Wirtes auf einen angepassten Krankheitserreger wird „Effektor getriggerte Immunität“ (ETI) genannt. Sie wird durch intrazelluläre Immunrezeptoren vermittelt, die einen spezifischen Virulenzfaktor des Krankheitserregers (Effektor) erkennen. Die meisten der bisher charakterisierten Immunrezeptoren gehören zu einer großen, sich schnell verändernden Familie von so genannten nucleotide-binding/leucine-rich repeat (NB-LRR)-Proteinen, die als molekulare Schalter fungieren [1]. Bei Pflanzen führt die Erkennungsreaktion zwischen Effektor und Rezeptor zu einer schnellen Änderung beim Ablesen der notwendigen Gene. Dieser Prozess wird meistens zusätzlich von einem programmierten Zelltod an der Infektionsstelle begleitet. Durch die zellulären Veränderungen werden alle nicht befallenen Teile der Pflanze gewarnt, sodass sie gegen weitere mögliche Angriffe gewappnet sind. Man spricht von systemischer Resistenz. Strikte Kontrollen müssen dabei verhindern, dass es nicht zu einer Autoimmunität kommt (Abb. 1).
Wie aus Aktivierung eine Abwehrreaktion wird
Obwohl man den Mechanismus der NB-LRR-Rezeptoraktivierung immer besser versteht, ist noch wenig darüber bekannt, wie daraus eine Abwehrreaktion entsteht. Einige Erkenntnisse konnten aus der durch das bakterielle Effektorprotein AvrRps4 angestoßenen Signalkette gewonnen werden. Bei Infektion wird AvrRps4 ins Innere der Blattzellen geschleust, um die Abwehr der Pflanzen zu unterbinden. Im Gegenzug versuchen zwei kooperierende NB-LRR-Immunrezeptoren - und zwar die Proteine RPS4 und RRS1 – die Wirkung des AvrRps4-Proteins zu bremsen [1, 2]. Beide Rezeptoren gehören zu einer Unterklasse von NB-LRR-Proteinen mit einer speziellen für die Signalübertragung zuständigen Domäne am N-Terminus der Polypeptidkette. Diese Region wird als „TIR-Domäne“ bezeichnet. Außerdem besitzt RRS1 eine DNA-Erkennungsdomäne am C-Terminus, mit der es an bestimmte Regionen der pflanzlichen DNA binden kann. Dieses Merkmal - und der Befund, dass sich RPS4 im Zellkern der Pflanzenzelle ansammeln muss, damit eine Resistenz ausgelöst werden kann [3] - macht deutlich, dass der Zellkern ein wichtiger Ort für die Perzeption und / oder Signalweiterleitung bei der Resistenzreaktion ist. Auch andersartige Immunrezeptoren müssen sich für eine Resistenzreaktion im Zellkern ansammeln, und außerdem weiß man, dass es einige pathogene Effektoren auf den Transkriptionsapparat des Wirtes abgesehen haben [2].
Ein weiterer Hauptakteur der angeborenen Immunabwehr ist das mobile, im Zellkern und im Zytoplasma vorhandene Protein EDS1. Dieses Protein bildet mit seinen beiden Partnerproteinen PAD4 und SAG101 unterschiedliche funktionale Komplexe in den beiden Zellkompartimenten [4, 5]. Eine Aufgabe von EDS1 besteht darin, bei der basalen Immunität gegenüber angepassten virulenten Krankheitserregern als positiver Regulator aufzutreten. Das erklärt möglicherweise auch, warum EDS1 und PAD4 in einkeimblättrigen Pflanzen vorkommen, die keine TIR-NB-LRR-Gene haben. In zweikeimblättrigen Pflanzen mit ihrem großen Repertoire an TIR-NB-LRR-Rezeptoren ist EDS1 auch für die ETI wichtig, die durch die TIR-NB-LRR-Rezeptoren vermittelt wird. Interessanterweise ist die Interaktion zwischen EDS1 und PAD4 nicht für eine lokale ETI notwendig, was wiederum dafür spricht, dass verschiedene molekulare Konfigurationen von EDS1 den unterschiedlichen Ausgang der Abwehr kontrollieren [4, 5]. EDS1 stellt damit offensichtlich eine Verbindung her zwischen der TIR-NB-LRR-Rezeptor-Erkennung von spezifischen pathogenen Effektoren und einem vielleicht sehr urtümlichen, weniger effektiven basalen Immunsystem.
Die Rolle von AvrRps4

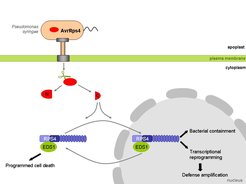
Ein anderer interessanter Mitspieler der RPS4/RRS1 Resistenz-Signalkaskade ist das AvrRps4-Protein selbst. Nachdem es in die Zelle gelangt ist, wandert sein C-Terminus zu der dem Zytoplasma zugewandten Seite von Endomembranen, wo der überwiegende Teil des zellulären RPS4-Pools lagert [3, 6]. Ein kleinerer Anteil von AvrRps4 gelangt auch in den Zellkern, wo das Protein mit EDS1 interagieren kann. Außerdem kann EDS1 auch Komplexe mit RPS4 im Zellkern bilden. Nach dem derzeitigen Kenntnisstand greift AvrRps4 das EDS1 Protein an, sobald es versucht, die basale Resistenz auszuschalten [6]. Dies wird durch die Bildung eines RPS4-EDS1-Komplexes unterbrochen, der dann eine ETI auslöst.
Wichtig ist, dass AvrRps4 keine angemessene Immunreaktion in Gang setzen kann, wenn es nur im Zyptoplasma vorkommt. Allerdings kann es dann den Tod von Wirtszellen herbeiführen [6]. Andererseits kann eine Ausbreitung der Bakterien begrenzt werden, wenn man AvrRps4 dazu zwingt, nur im Zellkern zu bleiben - dann unterbleibt allerdings der Zelltod [6]. Deshalb können Prozesse im Zellkern, die entscheidend für die Eingrenzung des bakteriellen Befalls sind, von denjenigen Prozessen im Zytoplasma unterschieden werden, die zu einem programmierten Zelltod führen - die aber wiederum nicht für die Begrenzung der Erkrankung nötig sind (Abb. 2). Vermutlich ist auch eine „Absprache“ zwischen den AvrRps4- und EDS1-Proteingehalten im Zellkern und im Zyptoplasma sowie deren Mobilität in der Zelle nötig, damit die für die systemische Resistenz nötige Verstärkung der Transkription im Wirt erreicht werden kann [6]. Um die verschiedenen Zweige der Immunabwehr zu bedienen, ist vermutlich zudem ein Hin und Herwandern von EDS1- Komplexen zwischen Kern und Zyptoplasma notwendig.
Als entscheidend für die Ausprägung einer effektiven und ausbalancierten Immunreaktion der Wirtspflanze zeichnet sich also die Kommunikation und dynamische Interaktion zwischen den verschiedenen Zellkompartimenten ab. Das Verstehen dieser Interaktionen wird zukünftiger Schwerpunkt der Forschung sein.